颜色是光线与感光系统互动的结果。人眼通过三种视锥细胞将连续光谱转换为三个神经信号,这解释了为何三原色能模拟色彩,并产生了同色异谱现象。
光谱分布 (SPD)
在理解计算机如何模拟颜色之前,必须澄清一个根本的物理事实:颜色并非物体固有的属性,而是光线与特定感光系统(如人眼、相机传感器)相互作用的产物。一个物体之所以呈现红色,并非因为它“是”红色,而是因为它吸收了可见光中除红光波段外的其他光线,并将红光反射到了我们的眼睛中。
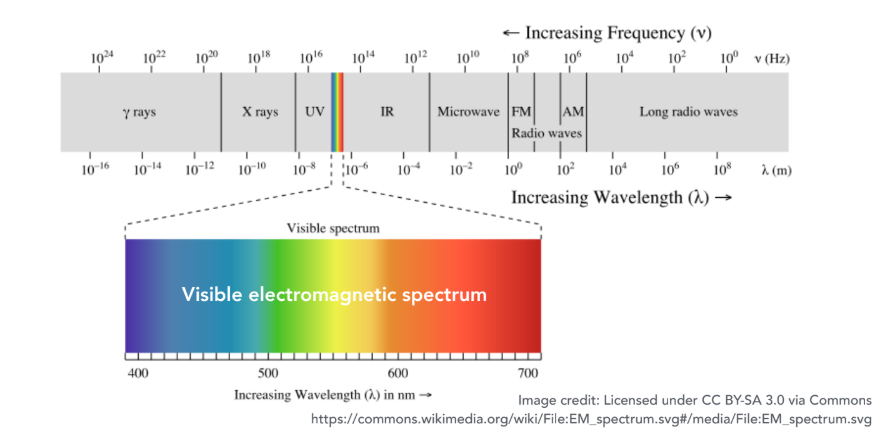
可见光谱 (Visible Spectrum):决定我们看到何种颜色的关键,是光的可见光谱。这是电磁波谱中波长介于 400纳米(紫光)到 700纳米(红光) 之间的一段。波长低于400nm的紫外线、高于700nm的红外线,人眼都无法直接感知。因此,在绝大多数游戏、影视渲染等视觉应用中,我们仅需模拟和计算这一狭小波长范围内的光线行为,这极大地简化了问题。
光谱功率分布 (SPD):为了精确描述一束光的颜色构成,我们使用光谱功率分布(Spectral Power Distribution, SPD) 这一核心物理量。SPD 描述了光在每个波长(λ)上携带的能量(或功率)。
它可以看作是一个函数:横坐标是波长 λ,纵坐标是该波长上的辐射能量。一束纯净的单色激光的 SPD 是一个尖锐的尖峰,而日常的太阳光或白炽灯光则是在整个可见光谱上都有广泛分布。SPD 是定义一种光最基础、最完整的物理方式。
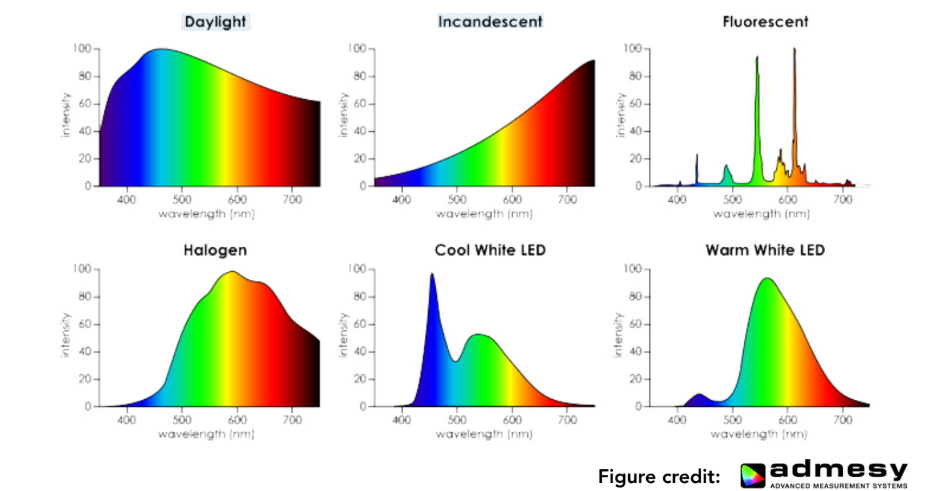
Tip SPD 就像是光的“成分配方表”。日光的配方比较均匀,而白炽灯的配方里“长波(红色)”比例更高 。
光线还具有线性性质:不同光线的能量(其 SPD)是线性可叠加的。
当两束或多束光交汇时,它们在空间任意一点上产生的总效果,就是各自 SPD 在该点的直接向量加法。这一原理是全局光照、多光源渲染、以及所有基于线性方程的光照模型(如辐射度、路径追踪)能够成立并正确计算的物理基础。正是基于这种线性叠加性,我们才能将复杂的光照场景分解为多个独立光源的贡献,分别计算后再合成最终图像。
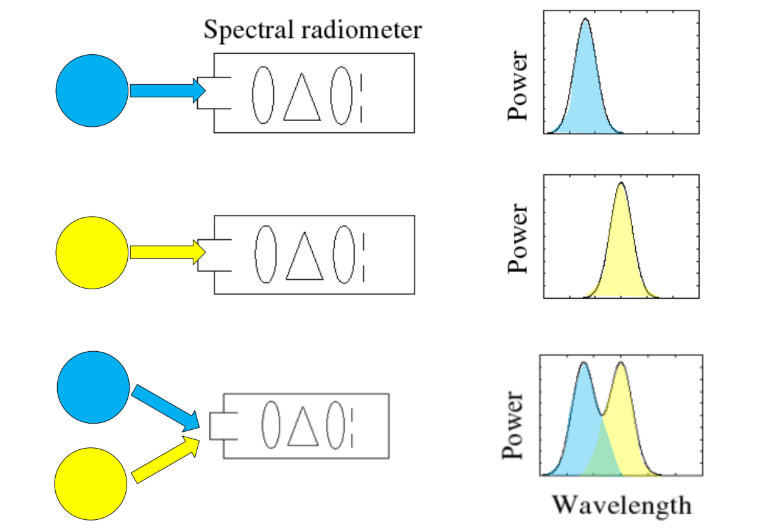
生物过滤网:大脑收到的三路信号
虽然自然界中的光在物理上拥有连续且近乎无限维的光谱分布,但人类之所以能用有限的“三原色”模拟和感知丰富的色彩世界,其根本原因在于我们视觉系统的生物学构造本身就是一个强大的“三通道生物滤波器”。
人眼包含两种功能不同的感光细胞:视杆细胞 负责在极暗环境下感知明暗,提供单色的夜视能力,但无法分辨颜色;而视锥细胞 则是在正常光照下产生色觉的主力。
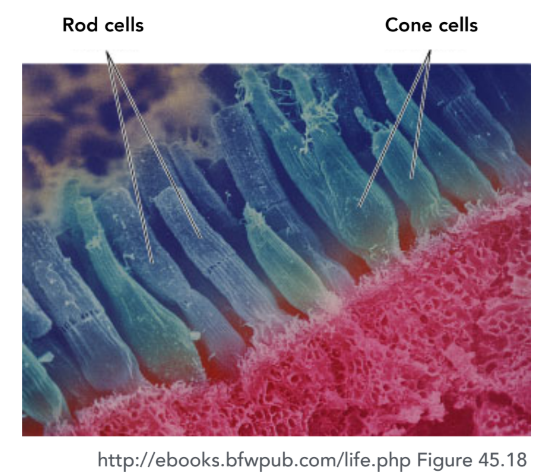
决定我们颜色视觉的关键在于,视锥细胞并非只有一种,而是主要分为三种,它们各自对不同波长的光具有最高的敏感度,但响应范围有所重叠:
- S-视锥细胞:对短波长(约 400-500nm,蓝色波段)最为敏感。
- M-视锥细胞:对中波长(约 450-630nm,绿色波段)最为敏感。
- L-视锥细胞:对长波长(约 500-700nm,红色波段)最为敏感。
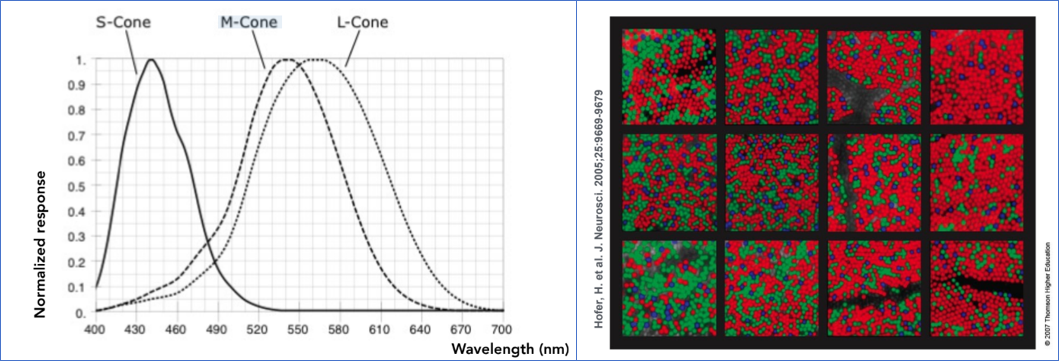
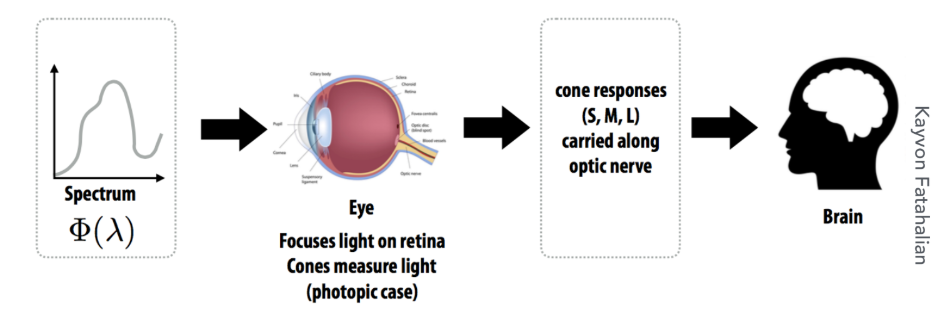
当我们观察一种光时,其完整的光谱功率分布 s(λ)并不会被大脑直接接收。相反,光首先经过这三种视锥细胞“过滤”。每种细胞的响应强度,并非简单地取决于其最敏感波长的能量,而是其自身的光谱敏感度曲线 r(λ)与入射光谱 s(λ)在整个可见光范围内的积分结果。即,大脑最终收到的信号是三个标量值:
$$ S = \int r_S(\lambda)s(\lambda)d\lambda $$$$ M = \int r_M(\lambda)s(\lambda)d\lambda $$$$ L = \int r_L(\lambda)s(\lambda)d\lambda $$这一生物学过程构成了三原色理论的生理学基础。 这意味着,无论自然界中光的物理光谱多么千变万化(是连续、无穷维的函数),进入人类视觉系统后,都被“压缩”或“编码”成了仅仅三个数值 (S,M,L)。因此,只要我们能找到三种特定的基础光源(即“三原色”),并通过调节它们的强度比例混合出某种光,使得这种混合光在三种视锥细胞中激发的 (S,M,L)响应与目标自然光相同,那么在人眼看来,这两种物理光谱完全不同的光,其颜色就是完全一样的。
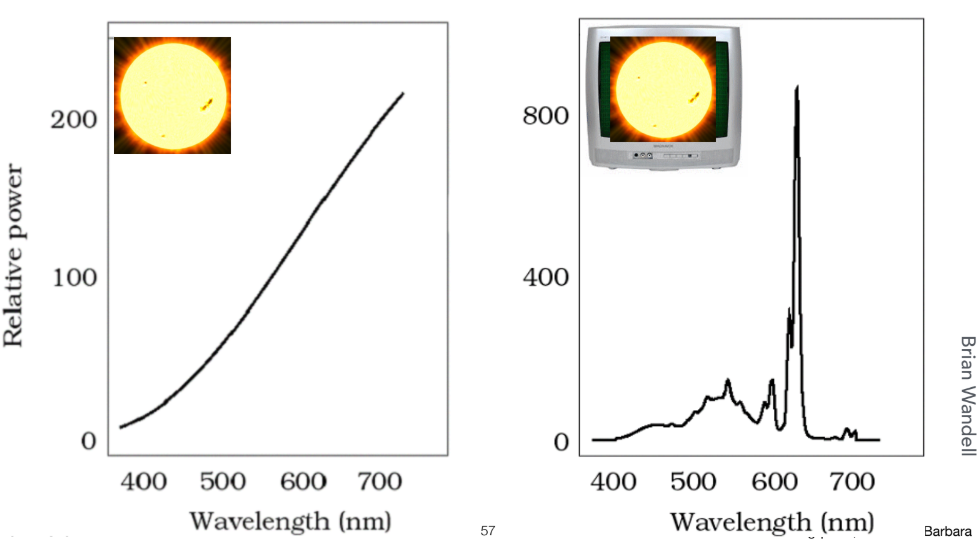
同色异谱 (Metamerism)
同色异谱 是颜色科学和图形学中一个极其精妙的现象,也是现代显示技术能够成功“欺骗”人眼的生理学基础。它描述了一种特殊情况:两种在物理光谱构成上完全不同的光,却能给人眼带来完全相同的颜色感知。
其数学原理直接源于人眼三色视觉的生物模型。假设有两种光,它们拥有不同的光谱功率分布 s1(λ)和 s2(λ)。当它们分别经过人眼的三种视锥细胞的敏感度曲线 rS(λ),rM(λ),rL(λ)进行“滤波”和积分后,如果产生的三刺激值 (S,M,L)完全相同,即:
$$ ∫rS(λ)s1(λ)dλ=∫rS(λ)s2(λ)dλ $$$$ ∫rM (λ)s1(λ)dλ=∫rM(λ)s2(λ)dλ $$$$ ∫rL (λ)s1 (λ)dλ=∫rL (λ)s2 (λ)dλ $$那么,尽管 $s1(λ)!=s2(λ)$,但是我们的大脑无法区分它们,会认为我们看到的是同一种颜色。
例如,显示器上的“白色”是由红、绿、蓝三个像素点以特定强度混合发出的光,其光谱是三个狭窄的尖峰。而自然界的日光是一种近乎连续的全光谱白光。这两种光的物理光谱分布天差地别,但通过精心设计的红、绿、蓝三原色强度配比,显示器可以模拟出与日光相同的 (S,M,L)三刺激值,从而让人眼产生“看到了日光”的错觉。因此,显示器不需要、也不可能复刻太阳的完整光谱,它只需巧妙地利用同色异谱效应,通过有限的三种光“凑”出相同的生物神经信号即可。
数学标准:从 RGB 到 XYZ
RGB 色彩匹配实验
为了将颜色感知量化,科学家设计了经典的色彩匹配实验。
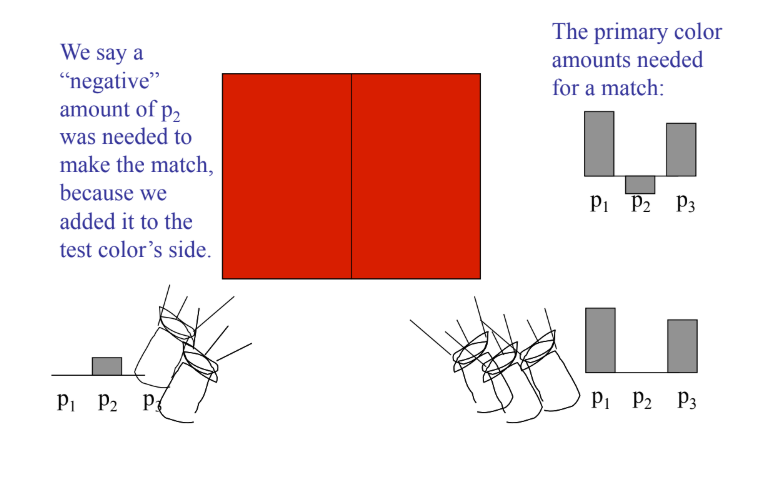
实验让观察者使用三束特定波长的单色光(通常为红、绿、蓝)进行混合,尝试匹配目标波长的单色光。
实验发现,大多数颜色可以通过调整三原色的比例来匹配,但某些颜色(如某些纯青色)无法通过简单相加获得。为了达到视觉匹配,有时必须将一部分红光加到目标光一侧,这在数学上相当于给红光赋予了负的权重。这说明,基于真实物理三原色的颜色系统在数学上存在负值,不利于计算和标准化。
CIE XYZ 空间:万能坐标
为了消除负值并建立一个普适、标准的颜色表示系统,国际照明委员会(CIE)在1931年定义了三个虚构的原色 X、Y、Z,对应三条经过精心设计的颜色匹配函数。这个系统的设计目标明确:
- Y 表示亮度:Y 通道被设计为与人类的明度感知直接对应,专门用于表示明暗程度。
- 无负值:所有可见光谱色在 XYZ 空间中的坐标值均为非负,消除了计算中的负权重问题。
- 全覆盖:X、Y、Z 构成的色域完全覆盖了所有人类可见的颜色。
由此,任何物理光谱都可以通过颜色匹配函数积分,转换为唯一的 XYZ 三刺激值,构成了颜色科学与工业的绝对基准。
色度图与色域
为了直观地研究颜色本身(排除亮度影响),我们通过归一化得到色度坐标:
$$ x = \frac{X}{X+Y+Z}, y = \frac{Y}{X+Y+Z} $$以 (x,y)为坐标绘制的图形,就是著名的 CIE 1931 色度图,其形状类似马蹄形(或舌形)。马蹄形边界线代表最纯净的单色光(光谱轨迹),内部是所有可能的可见颜色。
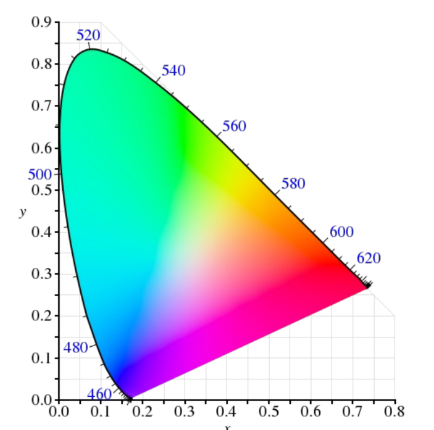
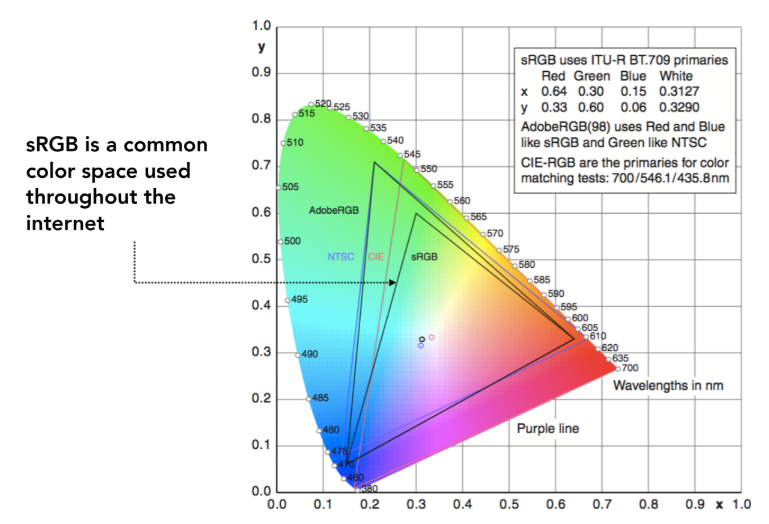
任何实际的显示设备(如显示器、打印机)都无法再现所有可见颜色,其能再现的颜色范围称为色域。在色度图中,一个设备的色域通常表示为一个三角形(如 sRGB、Adobe RGB、DCI-P3),三角形的顶点对应该设备三原色的色度坐标。三角形面积越大,代表该设备的色域越广,能显示的颜色越丰富。
感知均匀的颜色空间
HSV/HSB 空间
HSV(色相、饱和度、明度)是面向艺术家和设计师的直观颜色模型。它将颜色组织为:
- 色调 (Hue):围绕色相环的角度,表示“是什么颜色”(红、黄、绿等)。
- 饱和度 (Saturation):从中心(灰色)到边缘(纯色)的径向距离,表示颜色的鲜艳程度。
- 明度 (Value/Brightness):从黑到白的轴向位置,表示颜色的明暗。 这种模型更符合人类对颜色的直观描述习惯,便于进行选色和色彩调整。

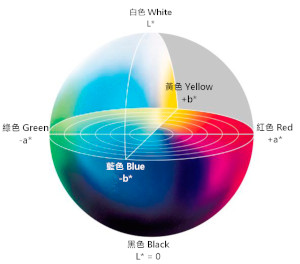
CIELAB 空间
虽然 XYZ 空间是标准,但它并非感知均匀的——即图中两点间的几何距离,并不对应人眼感知到的颜色差异大小。为了解决这个问题,CIE 推出了 CIELAB (L*a*b)* 颜色空间。
- L*:明度轴,从 0(黑)到 100(白)。
- a*:红-绿对立轴:正值偏红,负值偏绿。
- b*:黄-蓝对立轴:正值偏黄,负值偏蓝。 CIELAB 在设计上力求做到,在空间中任意方向上相等的几何距离,对应大致相等的视觉感知差异,因此在色彩管理、色彩差异评估等领域至关重要。
CIELAB 在设计上力求做到,在空间中任意方向上相等的几何距离,对应大致相等的视觉感知差异,因此在色彩管理、色彩差异评估等领域至关重要
拮抗色理论与“红绿色”的不可能性
CIELAB 的设计背后,有深刻的生理学基础——拮抗色理论。该理论认为,人脑并非独立处理红、绿、蓝信号,而是将其编码为三组对立通道:红-绿、蓝-黄、黑-白(明度)。
这解释了为什么我们无法想象或感知到“红绿色”或“蓝黄色”。因为红色和绿色处于同一根神经编码轴的两端,一个颜色信号只能在此轴上表现为偏红或偏绿,或处于中间的某种平衡(如黄或灰),但不能同时向两个极端偏移。就像一个物体不能同时处于“极大”和“极小”的状态。这种拮抗处理是视觉系统提高效率、增强对比的重要方式,也决定了我们感知和描述颜色的根本框架。